















But I'm getting ahead of myself. I just read (don't know why, really) an article by Mike Riddle, President of the Creation Training Initiative:
Does Evolution Have a . . . Chance?
Here's the whole things with my comments in red.
One has only to contemplate the magnitude of this task to concede that the spontaneous generation of a living organism is impossible. Yet we are here—as a result, I believe, of spontaneous generation.1
—George Wald, Nobel Laureate
—George Wald, Nobel Laureate
In today’s culture, molecules-to-man evolution is being taught as a fact, even though it is known to “go against the odds.” But few realize the odds they are up against! And they are immense!
The Bible teaches that God is the Creator of all things (Genesis 1; Colossians 1:16; John 1:1–3; Revelation 4:11 ). While these passages rule out any possibility of Darwinian evolution, they do allow for variation within a created kind. But there is much opposition to what the Bible teaches. People holding to evolution would argue that random chance events, natural selection, and billions of years are sufficient to account for the universe and all life forms. The fact they they rule out evolution merely means that they are wrong. Evolution - including macroevolution - has been observed.
Do You Believe in “Magic”?
Most people recognize “magic” as an illusionary feat or trickery by sleight of hand. But how far are you willing to go to believe something can happen by “dumb luck” or chance? For example, if I were to role a die and have it come up six three times in a row, would you consider that lucky? How about if I rolled six ten times in a row? Now you might suspect that I am using some trickery or that the die is weighted. It is much more incredulous to believe chance as an explanation than the magic of creationism. (Also, [sic]).
How far are we willing to go to accept something as a chance occurrence or before we recognize that it was just an illusion? We can test this by measuring our credulity factor. Credulity is the willingness to believe something on little evidence.
Measuring Our Credulity Factor against Evolution
Evolutionists state that life originated by natural processes about 3.8 billion years ago. Is there any evidence for this happening? Freeman Dyson, theoretical physicist, mathematician, and member of the U.S. National Academy of Sciences states:
Concerning the origin of life itself, the watershed between chemistry and biology, the transition between lifeless chemical activity and organized biological metabolism, there is no direct evidence at all. The crucial transition from disorder to order left behind no observable traces.2
Since the origin of life has never been observed, this is a major hurdle! Yes, true. It is a darned annoying fact that we cannot directly observe anything that happened in the past. If only we could directly observe murderers in the act, then detective work would be much easier. We are left with the question, “Is the origin of life by naturalistic processes possible?” This can, in part, be tested by examining two areas:
- The success of scientists in creating life or the components of a living cell.
- The probability that such an event could occur.
The Structural Unit of Living Organisms—The Cell
Cells are made up of thousands of components. One of these components is protein. Proteins are large molecules made up of a chain of amino acids. In order to get a protein useful for life, the correct amino acids must be linked together in the right order. There are of course many different ways to put together proteins that are useful for life. How easy is this and does it happen naturally? It turns out that this is not an easy process. No, not if your "process" is random chance with nothing else. There are large hurdles that evolutionary processes must overcome in order to build a biological protein.
Protein molecules contain very specific arrangements of amino acids. Even one missing or incorrect amino acid can lead to problems with the protein’s function. Yes, some amino acid changes will mess with protein function, but many changes are neutral and do not change protein function.
Making Mathematics Painless
Before applying mathematics and probability to the origin of life, we need to consider seven parameters that will affect the formation of a single protein.
First, there are over 300 different types of amino acids. However, only 20 different amino acids are used in life. This means that in order to have life, the selection process for building proteins must be very discriminating. But it didn't necessarily have to be this discriminating in the beginning.
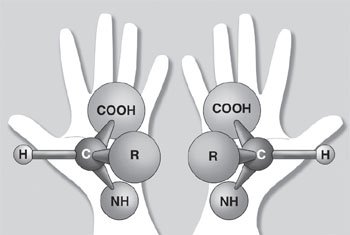
Second, each type of amino acid molecule comes in two shapes commonly referred to as left-handed and right-handed forms. Only left-handed amino acids are used in biological proteins; however, the natural tendency is for left- and right-handed amino acid molecules to bond indiscriminately.
Third, the various left-handed amino acids must bond in the correct order or the protein will not function properly.3 Again, there is not one correct protein, but a lot of variation, and proteins that doesn't work for one thing can work for another.
Fourth, if there was a pond of chemicals (“primordial soup”), it would have been diluted with many of the wrong types of amino acids and other chemicals available for bonding, making the proper amino acids no longer usable. This means there would have been fewer of the required amino acids used to build a biological protein. But there could have been enough. Plus, the twenty that are currently used could have been a function of those being to most abundant ones.
Fifth, amino acids require an energy source for bonding.4 Raw energy from the sun needs to be captured and converted into usable energy. Where did the energy converter come from? It would require energy to build this biological machine. However, before this energy converter can capture raw energy, it needs an energy source to build it—a catch-22 situation.5 See the video below.
Sixth, proteins without the protection of the cell membrane would disintegrate in water (hydrolysis), disintegrate in an atmosphere containing oxygen, and disintegrate due to the ultraviolet rays of the sun if there was no oxygen present to form the protective ozone layer.6
Seventh, natural selection cannot be invoked at the pre-biotic level. The first living cell must be in place before natural selection can function. No, selection works on anything that replicates. Self-replicating molecules like ribozymes are used in laboratory experiments. They are affected by natural selection.
Considering all seven of these hurdles, how probable is it that a single protein could have evolved from a pool of chemicals? Probability outcomes are measured with a value ranging from zero through one. The less likely an event will happen, the smaller the value (closer to zero). The more likely an event will occur, the larger the value (closer to one). Wow, talk about dumbing it down! If you know nothing about the natural processes that are involved, then it does seem very unlikely. But do watch this video to learn one or two things about those processes:
Let’s practice this using a coin. What are the chances of getting a heads when we flip a penny? The answer is 50 percent, or one chance in two (written 1/2). What is the chance of getting two heads in a row? Since each toss is 1/2 we can multiply each occurrence to get the final probability. This would be 1/2 x 1/2 which would equal 1/4 (or one chance in four). Now let’s use some bigger numbers.
When we flip a coin we have two possible outcomes, heads or tails. In this problem, we want to calculate the probability of getting all heads every time we flip a coin. We can use this exercise to test our credulity factor. How many heads in a row are we willing to accept as a chance occurrence? At what point would we suspect an illusion or some form of magic (trickery)? We wouldn't expect magic. Ever. Only godbots do that. We would instead expect some other natural process being involved.
The objective of using probabilities is to demonstrate the probability or chance of getting a certain result. On average, how many times and how often will we need to flip the coin to achieve 100 heads in a row? Over 300 million times a second for over one quadrillion years! If you could only do one trial at a time, then that would take a long time. But if you can do many at the same time in parallel, then you could get one hundred heads very quickly. If we could run just a billion such trials in parallel, then it would only take a million years, which is not long on geological time-scales. (Also, that number is slightly wrong. Only a little over 40 million times per second is needed for a quadrillion (1015) years. - My math is better than yours so I win!!! ;P)
The chances of getting all heads 100 times in a row is similar to the chance of getting 100 left-handed amino acids to form a biological protein. Proteins range in size from about 50 to over 30,000 amino acids. To get a small protein of 100 left-handed amino acids from an equal mixture of left- and right-handed amino acids, the probability would then be 1030 or 1 followed by 30 zeros (1,000,000,000,000,000,000,000,000,000,000). But but but, this is assuming that the process is random (again, it isn't - see the video above). How believable (credulity factor) is it that this could happen by random chance? Also, consider that this has never been observed! We all agree that it hasn't been observed, but we all agree that things that haven't been observed have taken place, right? Like a fallen tree in the forest is assumed to have fallen, even though no one were there to observe it. Chance protein formation has always been accepted as a matter of faith by evolutionists. No, not chance formation. Again, again, see the video. You are ignoring the natural processes that can explain these things.
| Number of desired heads in a row | Probability | Number of flips | Credulity factor (chance) |
| 1 | 1/2 have | 2 | Yes / No |
| 2 | 1/4 (1/22) | 4 | Yes / No |
| 3 | 1/8 (1/23) | 8 | Yes / No |
| 4 | 1/16 (1/24) | 16 | Yes / No |
| 5 | 1/32 (1/25) | 32 | Yes / No |
| 8 | 1/256 (1/28) | 256 | Yes / No |
| 10 | 1/1024 (1/210) | 1024 | Yes / No |
| 20 | 1/1,048,576 (1/220) | 1,048,576 | Yes / No |
| 100 | 1/1030(1/2100) | 1 followed by 30 zeros | Yes / No |
Ten is pretty good! We can work with ten. Not that we thereby admit that Riddle's puerile model here is the correct one (cause it isn't), but suppose to have a bunch of string of ten heads in a row, then those could be assembled together three at a time to make strings of 30 heads in a row.
But wait, there is more! This number, 1030, only measures the possibility of getting all left-handed amino acids. It does not say anything about their order. In our example, we have a chain of 100 amino acids. Each position can be occupied by any 1 of 20 different amino acids common to living things, and these must be in a specific order to form a functional protein. What is the probability that the correct amino acid will be placed in position number 1 of the chain? It will be 1/20. What is the probability that the first two positions will be correct? This can be calculated by multiplying the two probabilities together (1/20 x 1/20 = 1/202). Therefore, the probability of getting all 100 amino acids in the correct position would be 1/20 multiplied by itself 100 times or 1/20100 (this equates to 1/10130). This is 1 followed by 130 zeros! Which is not how proteins are thought to have formed. See the video above. This is like me saying that the process by which the Bible is written is by randomly stringing letters together. There are 3,566,480 letters in the bible (Bing it yourself), so with 26 different letters that gives a chance of one in 263566480. This is 1 followed by more than 5 million zeros! Therefore the Bible could not have been written by random chance. - Point here being that that is of course not the process by which the Bible was written, just as proteins of length 100 are not assembled by chance.

Large numbers can be hard to visualize or even comprehend. To put this in picture format we can use a smaller number 1021 (1 followed by 21 zeros). If we were to take 1021 silver dollars and lay them on the face of the earth; they would cover the entire land surface to a depth of 120 feet.7
Are there upper limits for which we can logically expect an event will not occur by random chance? The mathematician Emile Borel proposed 1/1050 as a universal probability bound. This means that any specified event beyond this value would be improbable and could not be attributed to chance.8 Repeat after me: scientists do not attribute random chance to the formation or proteins.
As we can see, the probability of getting a single small protein (1/10130) far exceeds this limit. Even if the protein can interchange amino acids at various positions (such as in the case of the protein cytochrome a),9 the resulting probability still exceeds the limit of 1/1050. So far we have only looked at the probability of getting a single small protein by random chance. What are the chances of getting all the proteins necessary for life? By chance? Negligible. Relevance...?
No matter how large the environment one considers, life cannot have had a random beginning . . . there are about two thousand enzymes, and the chance of obtaining them all in a random trial is only one part in (1020)2000 = 1040,000, an outrageously small probability that could not be faced even if the whole universe consisted of organic soup.10
Let our conclusion be that life did not have a random beginning (that is, completely random, as described here).
This number is so large (1 followed by 40,000 zeros) that it staggers the imagination how life could have evolved by natural, random processes. Yet, people continue to hold onto their belief that life did evolve by random chance (high credulity factor). Yes, staggering, I tell you. If you only rely on random processes, which scientists do not. Watch the video above!
Time is in fact the hero of the plot. . . . What we regard as impossible on the basis of human experience is meaningless here. Given so much time, the “impossible” becomes possible, the possible probable, and the probable virtually certain. One has only to wait: time itself performs the miracles.11

This statement attributes supernatural qualities to time! It also allows for anything to happen. This means we are no longer bound by the laws of science or any other natural limits. The statement thus becomes meaningless. You are the one not bound by the laws of science when you think the science says it is all random chance.
Tricks of the Trade
Since scientists have been unable to create life, they are forced to speculate through research and sometimes “sleight of hand” how it might have arrived on earth. Below are some of the tricks of the trade used to avoid the obvious—that God is the Creator of all things (Colossians 1:16). God or Allah or Odin or Zeus or Baal, or whatever. False dichotomy. Also, "speculate though research." *chortle* No, even if we understand natural processes that can create life, we can never know for sure how it actually happened, because all evidence of it has been erased. There are no fossils or anything else left from back then that we can take a direct look at. Too bad. But we can make very informed models by which we can understand abiogenesis. Sorry if this offends your religious sensibilities.
1. It happens naturally
“The formation of biological polymers from monomers is a function of the laws of chemistry and biochemistry, and these are decidedly not random.”12 This is a link to a great discussion on the probability of abiogenesis on TalkOrigins. It is also from 1998, and we have learned a lot since then. See, for example, the video above.
Explanation
This is an incorrect statement. I see nothing incorrect about it! Those laws are really not random!!! If it happens naturally, then why can’t scientists duplicate this in the lab? See video above. Amino acids do not spontaneously bond together to make proteins. First, it takes a source of energy to do this. The Sun or geothermal energy. Second, the natural tendency is to bond left- and right-handed amino acids, but life requires all left-handed amino acids. Third, they must be in the correct order or the protein will not function properly. Fourth, it requires the instructions of DNA to get the right amino acids. Where did DNA come from? Fifth, protein molecules tend to break down in the presence of oxygen or water. For answer to all of these, see the video above.
2. The deck of 52 cards
In a deck of 52 playing cards there are almost 1068 possible orderings of the cards. If we shuffle the deck we can conclude that the possible ordering of the cards having occurred in the order we got is 1 chance in 1068. This is certainly highly improbable, but we did come up with this exact order of cards. Therefore, no matter how low the probability, events can still occur and evolution is not mathematically impossible.
Explanation
In this example the math is correct but the interpretation is wrong. Your interpretation of your math is what's wrong here. If the arrangement had been specified beforehand, then the actual outcome would be surprising. By shuffling the cards, the probability is one that a sequence will occur. The fallacy is that the order is predicted after the fact. Your fallacy is that you assumed that there is just one correct protein, and is contains a hundred amino acids. That is false.
3. All the people
We are in a room of 100 people. What is the probability that all 100 people would be here in this room at this exact time? The probability is enormous, but yet we are all here.
Explanation
Two things are wrong with this reasoning. First, the people were not pre-specified. This is another example of an after-the-fact prediction. Second, each person made a decision to attend; therefore, this is not a chance gathering. This turns out to be a misunderstanding between a chance event and intelligent choice. Right! Just like proteins are not chance gatherings. There are natural non-random processes involved. I think you are getting it now.
4. Probability is not involved
Probability has nothing to do with evolution because evolution has no goal or objective.
Explanation
This statement disagrees with modern biology textbooks. Agreed. Probability does have something to do with it. I don't know where the quote in point 4 comes from (it isn't in the TalkOrigins article). It's just your probability calculations that have the wrong premises, namely ignoring lots of natural processes.
When there is more than one possible outcome and the outcome is not predetermined, probability can become a factor. In the case of evolution there is no pre-assigned chemical arrangement of amino acids to form a protein. Right again! (Yeah!) There are indeed not only one possible outcome, but many proteins that could work. Therefore, the formation of a biological protein is based on random chance. No, that really doesn't follow. I thought for a moment you were with us, but science lost you again. Scientists know today that it is only because of the instructions (information) in DNA that only left-handed amino acids are linked in the proper order.
Cells link amino acids together into proteins, but only according to instructions encoded in DNA and carried in RNA.13
Both creationists and evolutionists agree that DNA is essential for linking the correct amino acids in a chain to form a protein. The unanswered question is, “Where and how did DNA acquire the enormous amount of information (instructions) to form a protein?” There is no known natural explanation that can adequately explain the origin of life, or even a single protein. Yes there is. See the vid... The evolutionists are then left to rely on the odds (chance) that such a tremendous, improbable event occurred. No, there are other processes. zomg! Molecular biologist Michel Denton puts the event in perspective:
Is it really credible that random processes could have constructed a reality, the smallest element of which—a functional protein or gene—is complex beyond our own creative capacities, a reality which is the very antithesis of chance, which excels in every sense anything produced by the intelligence of man?14
But wait, there is still more!
The Human Body, Time, and Evolution
It is estimated that the human body is made up of 60 trillion cells (60,000,000,000,000).15 How long would it take to just assemble this many cells, one at a time and in no particular order at the rate of: What the fuck does this have to do with anything?!? Who thinks that the human body is assembled one cell at a time? Also, this doesn't seem to have anything to do with evolution, but development - a process that we can and have observed directly.
| One per second | 1.9 million years |
| One per minute | 114 million years |
| One per hour | 6.8 billion years |
These ages assume no mistakes! However, the evolutionary mechanism is based upon random errors (mistakes) in the DNA. Also included in assembling all the 60 trillion cells is that they have to make the right organs which all have to interact. Relevance?
The human body contains more than 40 billion capillaries extending over 25,000 miles, a heart that pumps over 100,000 times a day, red blood cells that transport oxygen to tissues, white blood cells that rush to identify enemy agents in the body and mark them for destruction, eyes and ears that are more complex than any man-made machine, a brain that contains over 100 trillion interconnections, plus many other parts such as the nervous system, skeleton, liver, lungs, skin, stomach, and kidneys. Relevance?
The complexity and dimensions of the human body are staggering. The probability of assembling 60 trillion cells that form specific organs that all work together to form a single human being in the evolutionary time scale of 3.8 billion years is a giant leap of faith. However, an all-knowing, all-powerful Creator has told us in His Word that He is the designer. That's not how anybody thinks the human body develops!
The hearing ear and the seeing eye, The Lord has made them both (Proverbs 20:12).
Every human body is a testimony to a purposeful Creator. As Malcolm Muggeridge said:
One of the peculiar sins of the twentieth century which we’ve developed to a very high level is the sin of credulity. It has been said that when human beings stop believing in God they believe in nothing. The truth is much worse: they believe in anything.16Nonsense! Both statements are false. I believe in many things, and God is not one of them.
Conclusion
Probability arguments can present a strong argument for the existence of a Creator God. The probability arguments presented here - even if they were based on sound assumptions, which they aren't - argues nothing for the existence of a Creator God. Certainly not any particular God. Maybe FSM. However, even when such evidence is presented to an evolutionist there is no guarantee that he or she will be persuaded. No, immature arguments like these persuade no scientists. Creationists, maybe. The real issue is not about evidence If you admit that you think it has nothing to do with evidence, why are you going through all these exercises in the first place?; it is a heart issue. As Christians we are called to have ready answers and break down strongholds that act as stumbling blocks to the unbeliever. It is the Holy Spirit that changes lives.
Your real creationist weapon is ignorance - something that you rely heavily on when calculating probabilities for protein formation by random chance alone.But sanctify the Lord God in your hearts, and always be ready to give a defense to everyone who asks you a reason for the hope that is in you, with meekness and fear (1 Peter 3:15).For the weapons of our warfare are not carnal but mighty in God for pulling down strongholds, casting down arguments and every high thing that exalts itself against the knowledge of God, bringing every thought into captivity to the obedience of Christ (2 Corinthians 10:4–5).
Footnotes
- George Wald [biochemist and winner of Noble Prize in Physiology or Medicine, 1967], “The Origin of Life,”Scientific American 191 no. 48 (1954): 46.
- Freeman Dyson, Origins of Life (New York, NY: Cambridge University Press, 1999), p. 36.
- “The order of the amino acids in a protein determines its function and whether indeed it will have a function at all.” Lee Spetner, Not By Chance (New York, NY: Judaica Press, 1997), p. 31.
- “The important fact that amino acids do not combine spontaneously, but require an input of energy, is a special problem.” Charles Thaxton, Walter Bradley, and Roger Olsen, The Mystery of Life’s Origin (Dallas, TX: Lewis and Stanley, 1992), p. 55.
- “A source of energy alone is not sufficient, however, to explain the origin or maintenance of living systems. The additional crucial factor is a means of converting this energy into the necessary useful work to build and maintain complex living systems.” Thaxton, Bradley, and Olsen, The Mystery of Life’s Origin, p. 124.
- “What we have then is a sort of ‘catch 22’ situation. If we have oxygen we have no organic compounds, but if we don’t have oxygen we have none either.” Michael Denton, Evolution: A Theory in Crisis (Bethesda, MD: Adler and Adler, 1985), p. 262.
- Peter Stoner, Science Speaks (Wheaton, IL: Van Kampen Press, 1952), p. 75.
- Emile Borel, Probabilities and Life (New York, NY: Dover, 1962), p. 28.
- A transport protein involved in the transfer of energy (electrons) within cells.
- Sir Fred Hoyle and Chandra Wickramasinghe, Evolution from Space (London: Dent, 1981), p. 148, 24.
- George Wald, “The Origin of Life,” p.48.
- Ian Musgrave, “Lies, Damned Lies, Statistics, and Probability of Abiogenesis Calculations,” TalkOrigins, www.talkorigins.org/faqs/abioprob/abioprob.html.
- G.B. Johnson, Biology: Visualizing Life (Austin, TX: Holt, Rinehart, and Winston, 1998), p. 193.
- Denton, Evolution: A Theory in Crisis, p. 342.
- Boyce Rensberger, Life Itself (New York, NY: Oxford University Press, 1996), p. 11.
- Malcolm Muggeridge, “An Eighth Deadly Sin,” Woman’s Hour radio broadcast, March 23, 1966. Quoted in Malcolm Muggeridge and Christopher Ralling, Muggeridge Through the Microphone: B.B.C. Radio and Television(London: British Broadcasting Corporation, 1967).









